分類について
目次
<タイトルをクリックすると該当箇所へジャンプします>
- 1. まりもの仲間の特徴
- 2. マリモ Aegagropila brownii (Dillwyn) Kützing 1854
- 3. モトスマリモ (甲府) Aegagropilopsis clavuligera (Grunow) Boedecker 2012
- 4. モトスマリモ (川崎) Aegagropilopsis clavuligera (Grunow) Boedecker 2012
- 5. タテヤママリモ Aegagropilopsis moravica (Dvořák) Z.-J.Zhao et G.-X.Liu 2015
- 6. アオミソウ Pithophora roettleri (Roth) Wittrock 1877
まりもの仲間の特徴
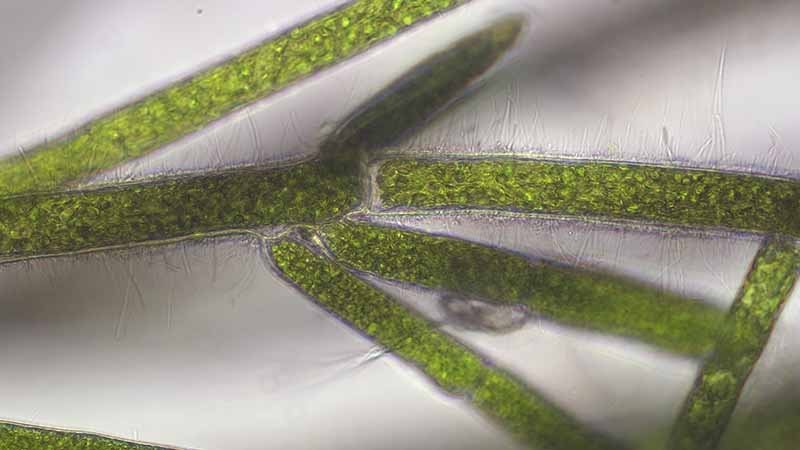
まりもの仲間は球状の形が特徴的で、「毬のような藻」つまり「まりも」と言われているわけですが、常に丸いというわけではありません。 石や岩にくっついて苔状または房状になったものや、水底でふわふわと漂うマット状のものもあります。これらをよく見ると、どれも緑色の細長い繊維状のものがたくさん集まっていることがわかります。 この繊維状のものを糸状体(しじょうたい)といいます。糸状体を顕微鏡で観察すると、円筒形の緑色の細胞が一列につながっていて、ところどころから枝分かれしています。細胞の太さや枝分かれの様子はまりもの種類によって違います。分枝は対になって出る(図1)、片側に偏っている(図2)、たくさん出る(図3)、垂直に出る(図4)、不規則に出る(図5)など様々です。また糸状体の一部の細胞が細長く伸びて他の糸状体に絡みついたり(図2、図6)、あるいは他の細胞より太くなって石や砂に付着したりすることがあります(図7)。これらを仮根(かこん)と言います。



(タテヤママリモ)



球形のまりもがバラバラになっても、小さな塊から再び大きく成長できます。また一本の糸状体や、糸状体から離れた枝からでも新しい個体に成長することができます。植物の挿し木と同じで栄養生殖と呼ばれます。
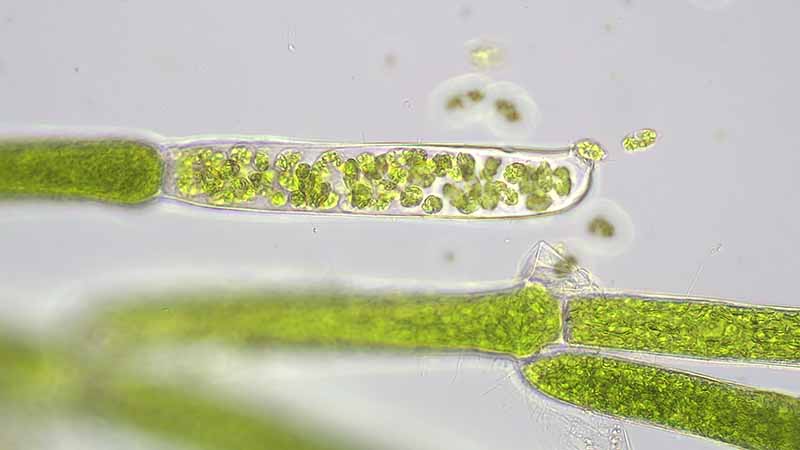
私たち動物も植物も、通常は細胞1個に核が1個ずつあります。しかし、まりもの仲間では1個の細胞に多数の核があります。次の世代をつくる仕組みも私たちとは違っています。モトスマリモでは何らかの条件によって枝の細胞が遊走子嚢という特殊な細胞に変わります。その中で細胞分裂が起こり、核を1個だけ持つ滴形の小さな細胞がたくさんできます(図8)。これを遊走子と言います。遊走子は光を感知できる眼点を1個と、先端に同じ長さの鞭毛を2本もっています。遊走子嚢の先端または側方に孔が1つ開き、そこから遊走子が1個ずつ出て泳ぎだします(図9)。遊走子は適当な場所に付着して伸長し、細胞分裂を繰り返して、親と同じような糸状体に成長します。遊走子嚢から遊走子が全部出てしまうと、遊走子嚢は透明になります(図10、図11)。タテヤママリモとマリモも同じように遊走子ができると考えられています。マリモの遊走子形成は非常に稀で、主に栄養生殖によって増えていると考えられてきましたが、夏場に遊走子を形成していることが分かってきました。また、従来見つかっていた2本鞭毛の遊走子に加えて4本鞭毛の遊走子も見つかりました。マリモは2本鞭毛の雄雌配偶子の接合による有性生殖を行っているようです(Umekawa et al. 2021)。この有性生殖がタテヤママリモやモトスマリモでも見つかるかどうかは今後の課題です。

中に遊走子が多数つまっている (モトスマリモ(甲府))

糸状体のあちこちに見られる (モトスマリモ(甲府))

遊走子嚢の先端には孔が一個ずつ開いている (モトスマリモ(川崎))
一方、アオミソウにはアキネートという生殖のための細胞ができますが(図12、図13、図14)、遊走子嚢や遊走子は知られていません。逆にアオミソウ以外のまりもの仲間ではアキネートは見つかっていません。そのため、アオミソウはまりもの仲間として扱われてこなかったのですが、遺伝的にも近縁で球形のまりもを作ることからここではまりもの仲間として扱っています。まりもの仲間の祖先は海に生育していたと考えられていて、その一群が汽水から淡水に侵入してきました。この一群をまりもの仲間として扱っています。

アキネートは濃緑色で、周囲の細胞は葉緑体が少なくて白色を呈する (アオミソウ)


アキネートという生殖のための細胞が糸状体の先端と中間の細胞の一部にできますが、遊走子嚢や遊走子は知られていません。アキネートは他の細胞より濃い緑色をしており、形は様々ですが、通常の細胞より短くやや太いので容易に見分けられます(図13)。アキネートができると、緑色が薄い細胞と濃い細胞が順番に並ぶので、肉眼で縞模様に見えます。アキネートから新しい細胞が発芽して、親と同じような糸状体にまで成長します。アキネートは元の糸状体から外れる場合と、そのままの位置から発芽する場合があります(図14)。
まりもの仲間の日本での生育地は次の通りです。マリモは火山性堰止湖(阿寒湖、富士五湖)、海跡湖(塘路湖、左京沼)、琵琶湖から見つかっています。タテヤママリモは湧水、河川、海跡湖、琵琶湖から見つかっています。琵琶湖や阿寒湖ではマリモとタテヤママリモが両方見つかっています。アオミソウは関東以南の田んぼなど様々な水域から報告されています。一方、モトスマリモは家庭の水槽からしか見つかっていません。2022年の私たちの論文が日本におけるモトスマリモの初報告ですが(Tuji & Niiyama 2022)、その後、日本各地の家庭の水槽に生育していることがわかりました。しかし、未だ野外では見つかっていません。最初に甲府から見つかったものと、次いで川崎から見つかったものは遺伝的にわずかな違いがあり、形態的にも異なりますので、一時的ですがモトスマリモ(甲府)、モトスマリモ(川崎)と分けて扱っています。
まりもの仲間のそれぞれの種類についての詳細については、各種類の解説をご覧ください。それぞれの解説で引用した文献はまとめて以下に示します。
引用文献
- Boedeker, C., Eggert, A., Immers, A. and Smets, E. 2010. Global decline of and threats to Aegagropila linnaei, with special reference to the lake ball habit. Bioscience 60: 187-198.
- Boedeker, C., O'Kelly, C. J., Star, W. and Frederik, L. 2012. Molecular phylogeny and taxonomy of the Aegagropila clade (Cladophorales, Ulvophyceae), including the description of Aegagropilopsis gen. nov. and Pseudocladophora gen. nov. Journal of Phycology 48: 808-825.
- Guiry, M.D. and Frödén, P. 2023. Aegagropila brownii (Dillwyn) Kützing is the correct name for Aegagropila linnaei Kützing, the type of Aegagropila Kützing (Pithophoraceae, Chlorophyta), widely known as “Marimo” or “Cladophora Balls”. Natulae Algarum 293: 1-7.
- Hanyuda, T., Wakana, I., Arai, S., Miyaji, K., Watano, Y. and Ueda, K. 2002. Phylogenetic relationships within Cladophorales(Ulvophyceae, Chlorophyta)inferred from 18SrRNA gene sequences, with special reference to Aegagropila linnaei. Journal of Phycology 38: 564-571.
- Hirose, H., Yamagishi, T. and Akiyama, M. 1977. Illustrations of the Japanese fresh-water algae. 933pp. Uchida Rokakuho, Tokyo.
- Hoek, C. van den. 1963. Revision of the European species of Cladophora. 248 pp. E.J. Brill, Leiden.
- Nakai, R., Wakana, I. and Niki, H. 2021. Internal microbial zonation during the massive growth of marimo. A lake ball of Aegafropila linnaei in Lake Akan. iScinece 24. 102720.
- Niiyama, Y. 1989. Morphology and classification of Cladophora aegagropila (L.) Rabenhorst (Cladophorales, Chlorophyta) in Japanese lakes. Phycologia 28: 70-76.
- Sakai, Y. 1964. The species of Cladophora from Japan and its vicinity. Scientific papers of the Institute of Algological Research, Faculty of Science, Hokkaido University 5: 1-104.
- Tuji, A and Niiyama, Y. 2022. First record of algal ball-forming Aegagropilopsis clavuligera (Cladophorales, Ulvophyceae) from Japan. Bulletin of National Museum of Nature and Science, Series B 48: 109-116.
- Tuji, A and Niiyama, Y. 2024. The second record of algal ball-forming Aegagropilopsis clavuligera (Ulvophyceae, Cladophorales) from Japan. Bulletin of National Museum of Nature and Science, Series B 50: 51-58.
- Umekawa, T., Wakana, I. and Ohara, M. 2021. Reproductive behavior and role in maintaining an aggregative form of the freshwater green alga Marimo, Aegagropila linnaei, in Lake Akan, Hokkaido, Japan. Aquatic Botany 168: 103309.
マリモ Aegagropila brownii (Dillwyn) Kützing 1854
表面がフェルト状のやや硬い球体をつくります。球の直径は数cmから最大30cm近くになりますが、大きなものは中空になっていて、中に泥や時にはミミズが入っていることもあります。マリモは丸くならず、湖底の石などに付着することも水底に漂って生育している場合もあります。暑さに弱く、成長はゆっくりです。糸状体は二又に次々に枝分かれを繰り返しながら長く伸びます。細胞は円筒形で、枝の先端は丸みを帯びています。糸状体の下方の細胞は中央や片側が膨らんでいることがあります。そのような細胞は細胞壁が厚くなっています。日本のマリモの糸状体の大きさは、主軸細胞が太さ約40-120µm、長さ約120-880µm、分枝の先端細胞が太さ約20-80µm、長さ約168-2000µmです(Niiyama1989)。
きれいなマリモ球体はどのようにできるのでしょうか。それにはマリモが生育している環境、つまり底質が泥か砂か礫か、水の流れは速いか、水深はどのくらいかなどが影響すると考えられています。付着したまま生育するものや、遊離した糸状体が絡み合って球状の塊まで大きくなることもあれば、さらに大きく成長した球形のマリモが壊れて小さな塊になり、再度きれいな球形になると考えられています。20cmを超す大きな球形に成長する際には、バクテリアが糊の役割を果たしているのではないかと考えられています(Nakai et al. 2021)。
これまでにマリモは日本では火山性堰止湖(阿寒湖、富士五湖)、海跡湖(塘路湖、左京沼など)、琵琶湖から見つかっています。ヨーロッパ各国の淡水湖や塩分濃度が0.6%以下の汽水域(バルト海、黒海、カスピ海)、ロシア、北米、オーストラリアなどからも報告されています(Hoek 1963)。しかし、世界各地で生育地と数が減っていると言われています(Boedeker et al. 2010)。特にきれいな球体マリモの減少は著しく、レッドリストに絶滅危惧種として掲載している国もあります。減少の理由は、湖沼の富栄養化や生育地の開発、乱獲などだと考えられます(Boedeker et al. 2010)。前述のようにマリモは成長が遅く、暑さに弱いという特徴があります。温暖化が進む現状では、きれいな球形のマリモが生き延びるのは至難の業と言えるでしょう。
日本でのマリモの発見は1889年に川上瀧彌が阿寒湖から報告したものが初めてで、Cladophora sauteri (Nees) Trevisanと同定されました。マリモの分類は主に形態的な特徴で行われ、阿寒湖以外の湖沼から報告されて色々な学名で呼ばれてきました(Sakai 1964; Hirose et al. 1977)。一方で欧州のマリモも日本のマリモも1種Cladophora aegagropila (L.) Rabenhorstであるという報告もあります(Hoek 1963; Niiyama 1989)。その後、遺伝子解析の結果が徐々に明らかになり、日本および世界各地で報告されているマリモはすべて同一種Aegagropila linnaei Kützing であることが確認されました。私たちもこの学名を使用してきたのですが、最近、より古い標本が見つかったことからマリモの学名はAegagropila brownii (Dillwyn) Kützingとすることが提案されました(Guiry & Frödén 2023)。私たちもここではこの新しい学名を採用します。
モトスマリモ(甲府) Aegagropilopsis clavuligera (Grunow) Boedecker 2012
柔らかく、表面が滑らかできれいな球体で、直径は数cmから最大5cmくらいです。糸状体はまっすぐ伸びて、たくさんの短い分枝がやや規則的に斜め上に向かって出ます。分枝は糸状体の片側に偏っていることが多いですが、対になって出ることもあります(図2)。 仮根が糸状体の下部や上部から出て、糸状体同士が絡み合っています(図2、図6)。そのためにきれいな球形が保たれているのかもしれません。細胞は円筒形で、枝の先端は丸みを帯びています。遊走子嚢の形成が頻繁に観察できます(図8、図10)。分枝の細胞が連続して遊走子嚢になる場合もあります。糸状体の主軸細胞の太さ約34-60µm、長さ約180-380µm、分枝細胞の太さ約21-39µm、長さ約133-590µm(Tuji & Niiyama 2022)。
日本では甲府市にお住いの方の水槽から初めて見つかりました(Tuji & Niiyama 2022)。その後、熱帯魚や水草を育てている日本各地の家庭の水槽からも見つかっていますが、川崎タイプのモトスマリモより少ないようです。野外からの報告はありません。遺伝子解析の結果、中国から報告されたものと近いことがわかっています(Tuji & Niiyama 2024)。野外でどのような生活をしているのか不明なことが多い種です。
モトスマリモ(川崎) Aegagropilopsis clavuligera (Grunow) Boedecker 2012
とても柔らかく、ふわふわした手触りの球形で、直径は最大5㎝くらいです。たくさんの房状に伸びた糸状体が集合して球体になっています。これまで水槽でしか見つかっていません。糸状体はまっすぐ伸びて、たくさんの短い分枝が規則的に並んで斜め上に向かって出ます(図1)。分枝は対になって左右に出ることが多いです。一か所から3、4本の分枝が出ることもあります(図3)。細胞は円筒形で、枝の先端は丸みを帯びています。遊走子嚢の形成が頻繁に観察できます(図9)。分枝の細胞が連続して遊走子嚢になる場合もあります(図11)。糸状体の主軸細胞の太さ約24-39µm、長さ約109-500µm、分枝細胞の太さ約15-24µm、長さ約87-587µm(Tuji & Niiyama 2024)。
日本では川崎市にお住いの方の屋内水槽から初めて見つかりました(Tuji & Niiyama 2024)。その後、熱帯魚や水草を育てている日本各地の家庭の水槽から見つかっていますが、野外からの報告はありません。本種の模式産地はスリランカで、淡水巻き貝の殻から採取されたものです(Boedecker 2012)。遺伝子解析の結果、日本で見つかった本種はオランダの熱帯水槽から見つかったものと近いことがわかりました(Tuji & Niiyama 2024)。本種も野外でどのような生活をしているのか、まだ不明な点が多い種です。モトスマリモ(川崎)は、モトスマリモ(甲府)に比べて糸状体同士の絡み合いが少なく、房状になります。また、モトスマリモ(川崎)の方が糸状体も細いため、両者は別種の可能性が高いと思われます。現在研究中です。
タテヤママリモ Aegagropilopsis moravica (Dvořák) Z.-J.Zhao et G.-X.Liu 2015
まりもは柔らかく、ややいびつな形です。最大3cmくらいですが、これより大きくなることがあります。これまで温帯の湧水で見つかっています。枝分かれは多様で、やや不規則に直角に出ることも、また短い枝や長い枝が二又に順次伸びることもあります。細胞は円筒形で、枝の先端は丸みを帯びています。糸状体の下方の細胞は太く、中央や片側が膨らむことがあります(図4)。また細長く伸びた仮根が糸状体のあちこちから生じ、別の分枝や隣接する糸状体や石などに絡みついているのが見られます。遊走子嚢の形成はモトスマリモほど頻繁には見られません。
本種は富山県立山町の湧水から初めて発見され、形態の比較と遺伝子解析が行われた結果、マリモとは全く別の種タテヤママリモであることが明らかになりました(Hanyuda et al. 2002)。
アオミソウ Pithophora roettleri (Roth) Wittrock 1877
ごわごわした手触りで、球形やマット状など様々な形になります。糸状体は他のまりもに比べて太く、枝分かれは少なく、枝の長さは様々で、先端はややとがっています。糸状体の主軸は太さ約50 -130µm、多くの場合80µm前後、長さは太さの約5-20倍、分枝細胞の太さ約35-70µmほどです(図5)。日本人の髪の毛は太さが50-150µm、平均約100µmと言われていますから、アオミソウの主軸の太さは毛髪ほどです。アオミソウはモトスマリモの約2倍の太さです。
アキネートという生殖のための細胞が糸状体の先端と中間の細胞の一部にできますが、遊走子嚢や遊走子は知られていません。アキネートは他の細胞より濃い緑色で、やや太く短いので容易に見分けられます(図12、図13)。アキネートができると、緑色が薄く細長い細胞と濃緑色でやや太い細胞が順番に並ぶので、肉眼でも縞模様が見えます。アキネートから新しい細胞が発芽して、親と同じような糸状体にまで成長します。アキネートは元の糸状体から外れる場合と、そのままの位置から発芽する場合があります(図14)。
アオミソウは暖かな気候を好み、暑さに強く、日本では関東以南で見られます。また熱帯や亜熱帯にも生育しています。
日本で最初にアオミソウが報告されたのは、1928年に東道太郎博士が植物研究雑誌に発表した「将に帰化植物トナラントスル淡水藻ノ一例」という論文でした。東博士はこの論文で、横浜市在住の日本人男性がサンフランシスコから取り寄せた熱帯魚に付着して「アクアリウム」中で繁茂した淡水藻をPithophora kewensis Wittrockと同定し、これにアオミソウという和名をつけて発表しました。同じ論文で、かつてPithophora zelleri (v. Marn.) Wittrockを千葉県浦安の水田で採集したけれども、すべての標本を1923年の関東大震災で焼失してしまったとも記しています。後にPithophora zelleriにはフシマダラという和名が付けられています(Hirose et al. 1977)。Pithophora属はかつて40種ほどが報告されていましたが、糸状体やアキネートの形態は非常に多様ではあるものの、形態と遺伝子解析の結果から、現在はすべて同じ種Pithophora roettleri (Roth) Wittrockであると考えられています(Boedeker et al. 2012)。最初に報告された和名はアオミソウですので、私たちはPithophora roettleriをアオミソウと呼んでいます。